Las proteínas que identifican y diferencian a las especies se originan en la información del DNA. En particular, el conocimiento del HLA se aplica hoy a los estudios epidemiológicos y la medicina preventiva, a la paleontología, la antropología o el estudio de las migraciones.
La extraordinaria variación de actividades fisiológicas que muestran los seres vivos y, además, la especificidad propia de cada especie, se deben principalmente a la presencia de las proteínas. Proteínas, por tanto que son capaces de exhibir actividades catalíticas, de transporte y almacenamiento, de soporte mecánico, como factores de crecimiento y diferenciación como receptores de membrana, de anticuerpos, de adhesión y de coordinación de movimientos. Proteínas que, de otro lado, revelan la especificidad como consecuencia de la ordenación de sus unidades constituyentes que, en número de veinte, se alternan de manera constante en cada especie. Secuencia conocida como estructura primaria.
Este lenguaje de los aminoácidos se mantiene constante a través de las sucesivas generaciones de la misma especie y es distinto, a su vez, en las diferentes especies de seres vivos. La semejanza de este lenguaje o, lo que es lo mismo, la analogía de estructuras primarias da cuenta del grado de aproximación entre las diferentes especies; entre el hombre y el chimpancé, entre el perro y el caballo, etc.
La interpretación de estos hechos, la constancia por un lado y el grado de semejanza por otro, se funda en el mismo hecho biológico fundamental: la especificidad estructural de las proteínas tiene su origen en la información propia y constante en cada especie y distinta entre las especies. Información que no es sino el DNA y, en particular, la secuencia que se establece al ordenarse las unidades constituyentes que, en número de cuatro, se alternan de forma constante en cada especie.
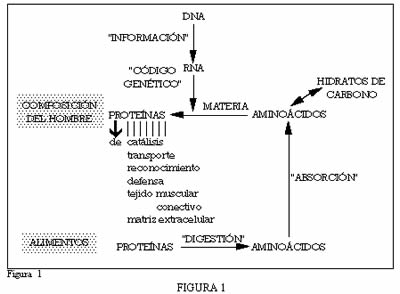
Este contenido informativo del DNA -y no su materia- es la causa de la especificidad mencionada que exhiben las proteínas. DNA presente en los genes de cada especie, que se expresa como proteínas específicas y ello se realiza a través de dos fenómenos principales: la transcripción de la información -paso de DNA a mRNA- y la traducción de la información -paso de la información del mRNA a la información de las proteínas-. Fenómenos que se resumen en la figura 1.
El DNA, con su clásica estructura en doble hélice, cumple en el núcleo de las células su doble función:
– La de servir de información a la especificidad de las proteínas.
– La de duplicarse durante el ciclo celular. Tras la interfase ocurre la división celular o mitosis, en cuya iniciación el DNA experimenta un curso de transformaciones que conduce a su condensación bajo la forma de cromosomas.
Los cromosomas difieren en el tamaño, la posición del centrómero y la presencia de regiones o bandas capaces de tinciones especiales. Los cromosomas, en esta fase, se hacen visibles al microscopio y su composición fotográfica da lugar al denominado cariotipo. El número de cromosomas es característico de cada especie, algunos de cuyos ejemplos aparecen en la tabla 1.
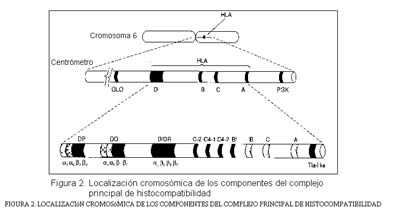
En la figura 2 se esquematiza la forma de un cromosoma, con el centrómero y los brazos cortos (p) y largos (q), en los que se alinean los genes. De la expresión de los genes (DNA) resulta la gran cantidad de proteínas específicas propias de cada especie.
En la tabla 2 se ha seleccionado una serie de localizaciones génicas en los cromosomas correspondientes; a la vez se mencionan las proteínas expresadas a expensas de los genes seleccionados. Dentro de estas localizaciones recogidas en la tabla 2, figura en el brazo corto del cromosoma 6 una situación de interés excepcional: la del llamado complejo principal de histocompatibilidad. Las propiedades principales de este complejo se refieren al polimorfismo, la codominancia y la transmisión conjunta.
Con estos antecedentes, ya se puede vislumbrar el interés biológico de este conjunto de genes. Interés que vamos a verlo extrapolado en cuanto a su participación en estudios geográficos, históricos, lingüisticos, antropológicos y epidemiológicos.
Poco antes de la II Guerra Mundial, en 1938, un genetista americano, Peter Gorer, demostró que los rechazos de los injertos de piel entre Ratones de cepas diferentes eran debidos a diferencias entre donadores y receptores a nivel de unas proteínas expresadas en el seno de las membranas celulares. Proteínas que fueron bautizadas como antígenos H-2; de la inicial H, de histocompatibilidad. Enseguida se constata que la aceptación o el rechazo de un transplante se encuentra bajo la dependencia de un proceso de naturaleza inmunológica, genéticamente determinado. Veinte años más tarde, el inmunólogo francés Dausset descubriría sobre la superficie de los linfocitos humanos unas moléculas homólogas que se denominaron antígenos HLA (Antígenos de los Leucocitos Humanos).
– La existencia en este sistema HLA de un extraordinario polimorfismo genético.
– Las moléculas HLA se encuentran bajo la dependencia de genes que pueden presentarse bajo formas diferentes o alelos, según los individuos considerados.
– Cada individuo expresa, al menos, seis tipos de estas moléculas, repartidas en dos grupos; las moléculas dichas de clase I (A,B,C) y las de clase II (DP,DR,DQ).
– Las variantes descritas hasta hace un año de cada uno de estos tipos de moléculas son, respectivamente: 23, 49, 8, 6, 3 y 25. Fenómeno conocido como polimorfismo, que puede dar lugar a cientos de millones de combinaciones polimórficas.
Los antígenos HLA ilustran como ningún otro marcador biológico la unicidad y la diversidad de los individuos. De otra manera, la probabilidad de identidad para todos los alelos entre dos individuos no relacionados es prácticamente nula; de aquí también la gran dificultad de encontrar un donador y un receptor del todo compatibles para el transplante de órganos.

La figura 2 recoge algunos datos acerca de la localización del complejo principal de histocompatibilidad.
Además de esta participación génica en cuanto a la definición de la individualidad del hombre, los antígenos HLA toman parte en algunos fenómenos de importancia tales como:
– La naturaleza y la intensidad de la respuesta inmune, en particulas la vinculada a los linfocitos T citotóxicos.
– Su correlación con el riesgo de padecimiento de ciertas enfermedades, sobre todo con aquellas de origen autoinmune.
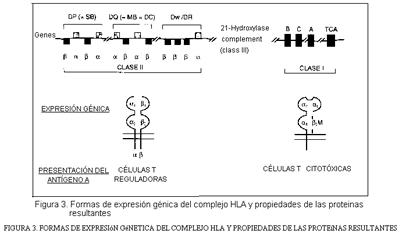
La conexión DNA-proteínas y la funcionalidad de éstas se esquematiza en la figura 3.
Veamos algunos detalles adicionales sobre las propiedades de este complejo.
En el hombre, los alelos A, B y C se expresan en todas las células del organismo bajo la forma de las llamadas proteínas de clase I (figura 3). Los alelos de la clase D no se expresan más que en ciertos tipos de células como macrófagos, espermatozoides, células epidérmicas, linfocitos E. Y ambos,antígenos de las clases I y II se expresan en las membranas celulares.
Por lo que se refiere a la complejidad del sistema, es obvio que ha tenido que darse sobre estos genes una presión particular para hacerlos diferentes y para mantener las diferencias. Dado que estas estructuras juegan un papel en el reconocimiento de sí mismo y que derivan de genes primitivos codificadores de moléculas de la superficie celular, se puede concebir la necesidad de esta diversidad cuando los seres vivos pasaran del estado unicelular o de una especie de conjunto de células idénticas al estado de organismo multicelular organizado cuyos tejidos hubieran de coexistir, e incluso cooperar. El mantenimiento de este polimorfismo es, indudablemente, una ventaja selectiva, a la que habrán de contribuir las funciones inmunes atribuidas a este complejo en el curso de la evolución.
El hecho de que los loci génicos A, B y C estén presentes en la superficie de prácticamente todas las células del organismo, sugiere una función biológica muy general. por el contrario, los productos de los loci D están muy restringidos a la superficie de ciertas células, ya mencionadas, esencialmente apoyada de competencia inmunológica. Circunstancia fundamental por lo que se refiere a la función de estos productos.
Los diferentes loci del complejo HLA se encuentran íntimamente ligados sobre el brazo corto del cromosoma 6. Hay, sin embargo, suficiente distancia entre ellos para que tengan lugar recombinaciones relativamente frecuentes entre ellos. Esta situación especial parece ser virtualmente única en la especie humana. Además, va acompañada de un fenómeno particular como es la asociación gamética preferencial entre alelos de los varios loci del complejo. Se dice en este caso que entre estos alelos se da un desequilibrio de ligamiento. O de otra manera, la magnitud de la desviación de las frecuencias génicas de los genes ligados respecto de lo esperado a partir de las frecuencias alélicas en la población, constituye el desequilibrio o asociación no al azar de los genes. Fenómeno que ha dado lugar a múltiples especulaciones: ¿Es una mera reminiscencia de combinaciones ancestrales, cuando, hace miles de años, las poblaciones estaban aisladas y que se manifestaron o incrementaron por las migraciones humanas?; ¿sería este desequilibrio una asociación preferencial seleccionada en virtud de las ventajas biológicas que ofreciese en un entorno determinado? Cabe también preguntarse si este desequilibrio de ligamiento es específico del complejo de histocompatibilidad o si, por el contrario, es un hecho más general con existencia en otras localizaciones del genoma.
Se trata, pues, de que el complejo HLA es el segmento del genoma humano mejor conocido y un excelente ejemplo de la relación de un producto con la secuencia del correspondiente gen.
Y ya se puede valorar el polimorfismo HLA y su desequilibrio de ligamiento como herramientas de gran utilidad en estudios antropológicos y epidemiológicos. A los primeros permite caracterizar poblaciones, discernir su origen y dibujar su historia genética. A los segundos comparar las fórmulas HLA con la susceptibilidad particular de poblaciones o grupos de poblaciones a ciertas enfermedades. y, quizá, en el futuro, reconstruir las grandes epidemias del pasado mediante la observación de las selecciones que operaron.
En la figura 4 aparece la disposición en las membranas celulares de los productos de la expresión de los genes del complejo.
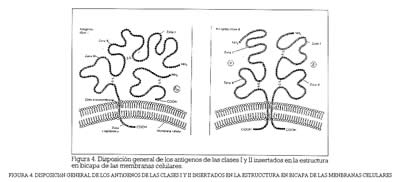
Los antígenos de la clase I constan de una larga cadena (a) con tres dominios principales, que interacciona con una molécula de ß2microglobulina. La cadena tiene su carboxilo terminal hacia la parte interior de la célula y el grupo amino terminal hacia la parte externa. Los antígenos de clase II poseen dos cadenas polipeptídicas, y ß, cada una de ellas con dos dominios, que atraviesan la bicapa lipídica de la membrana y situan los grupos carboxilo en el interior de la célula. El dominio III de la cadena y la cadena ß2 de microglobulina exhiben notables semejanzas con las estructuras de las inmunoglobulinas; lo que sugiere la posibilidad de un gen ancestral común. Por el contrario, las cadenas y ß de los antígenos de la clase II no muestran semejanza alguna con los productos de la clase I ni con las inmunoglobulinas. Todo parece como si los productos del complejo HLA fueran bipolares; es decir, derivados de dos genes muy distintos cuya función y origen se retrotraen muy lejanamente en la evolución de las especies.
El estudio de las estructuras polipeptídicas de ambas clases de productos, I y II, está permitiendo deducir datos acerca de las funciones específicas de cada una de las regiones de las moléculas.
Estas proteínas de las clases I y II desempeñan papeles distintos pero ambos fundamentales en la puesta en marcha de la comunicación inmunitaria; es el proceso conocido como el procesamiento de los antígenos. Según sea la naturaleza de los antígenos, así será el mecanismo de su procesamiento y así será la participación de las proteínas HLA, bien de la clase I o de la clase II.
Las proteínas de la clase I participan en el procesamiento de los antígenos citoplásmicos que se degradan en el seno del retículo endoplásmico por acción de un tipo singular de proteosomas celulares. Estos proteosomas están constituidos por el complejo LMP (low molecular mass polypeptide) que posee múltiples sitios catalíticos, de forma que al actuar sobre el sustrato celular produce simultáneamente un cierto número de péptidos (figura 5) con nueve aminoacidos.
Los péptidos así formados se transportan al lúmen del retículo endoplásmico a través de un heterodímero transportador, producto de expresión de los genes Tap-1 y Tap-2. En el lúmen, los péptidos interaccionan con las moléculas de clase I y gracias a un cambio conformacional se posibilita estas moleculas a la superficie celular.
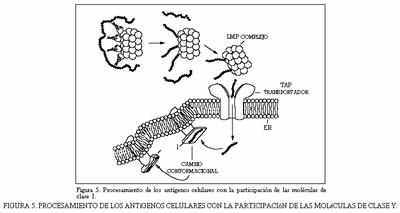
La dicotomía funcional entre las moléculas de clase I y las de la clase II se refleja también en la ruta intracelular seguida por ellas y por el antígeno. Así,las proteínas de la clase II tienen que ver con el procesamiento de los antígenos bacterianos y otras proteínas extracelulares.
Como ha quedado mencionado, la unidad funcional de las moléculas de la clase II consiste en un heterodímero ß. Durante la biosíntesis del heterodímero, éste permanece ligado a una tercera cadena, , cuya misión es la de proteger al dímero de su unión a péptidos indeseados durante su permanencia y migración en el retículo endoplásmico.
La cadena y se libera en el retículo trans-Golgi. Mientras tanto, los antígenos exteriores a las células se internan a través de etapas del tipo de gránulos recubiertos, endosomas primitivos y tardíos, hasta alcanzar los compartimientos MIIC en las células B. Los antígenos de esta manera aegradados interaccionan con las moléculas de clase II y los complejos clase II ( ß)-péptido se transportan a la superficie celular (figura 6).
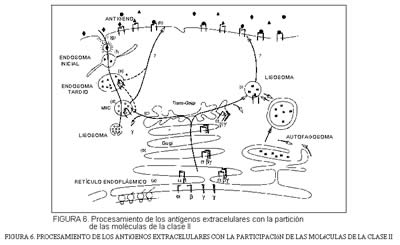
Así, pues, los antígenos intracelulares y extracelulares ofrecen oportunidades distintas al sistema inmune, tanto por lo que se refiere a su reconocimiento como a su adecuada respuesta; oportunidades diferentes aunque posean mecanismos que gozan de un cierto paralelismo.
Los péptidos derivados de los antígenos intracelulares se presentan por lo general a las células T CD8+ por las moléculas de clase I que se expresan virtualmente en todas las células; en tanto que los péptidos derivados de antígenos extracelulares se presentan por lo general a las células T CD4+ por las moléculas clase II presentes en células especializadas. Los detalles de estos mecanismos de la comunicación celular son objeto en la actualidad de intensos estudios.

Además del conjunto de circunstancias que subrayan el interés de los antígenos de histocompatibilidad, se da la condición ya mencionada de que ciertas enfermedades con características autoinmunes están asociadas a los antígenos HLA. Estas asociaciones son diferentes en las distintas poblaciones y su utilidad diagnóstica y epidemiológica exige, por tanto, definir dichas asociaciones en cada población. La tabla 3 reune algunos datos de este tipo de asociaciones.

Los antígenos HLA capacitan a las células del sistema inmune para llevar a cabo el reconocimiento de las proteínas extrañas invasoras del organismo. Las células T llevarán a cabo tal reconocimiento solamente cuando estén interaccionando con los antígenos HLA. Los genes HLA de la clase I gobiernan la actuación de las células T citotóxicas («killer») y los genes de la clase II gobiernan la de las células T auxiliares («helper»), inductoras de la producción de anticuerpos específicos por otras células del sistema inmune.
Las células T portadoras de receptores ß reconocen los péptidos antigénicos unidos a las proteínas de las clases I y II, codificadas en el complejo principal de histocompatibilidad. La estructura cristalográfica de estas proteínas ha mostrado la existencia de localizaciones a través de las que pueden unirse peptidos adecuados. Tras su síntesis, las proteínas de las clases I y II han adquirido una estructura terciaria parcial que se completa y logra la maduración conformacional por medio de la interacción con los péptidos.
Los antígenos intracelulares y extracelulares incitan diferentes respuestas del sistema inmune, tanto en términos de reconocimiento como de la respuesta apropiada. y, así, han evolucionado sistemas paralelos para llevar a cabo estas misiones. Los péptidos derivados de los antígenos intracelulares se presentan generalmente a las células T CD8+ por las moléculas de la clase I que se expresan virtualmente sobre todas las células; mientras que los péptidos derivados de los antígenos extracelulares son presentados a las células T CD4+ por las moléculas de la clase II, presentes tan sólo en células especializadas en la presentación de antígenos.
Aunque las proteínas clase I y clase II derivan del mismo conjunto de genes de región variable TCR, los procesos utilizados para originar los péptidos son diferentes. Las moléculas clase II obtienen sus péptidos predominantemente de fuentes extracelulares mediante endocitosis, con la degradación subsiguiente en lisosomas o endosomas en medio ácido; por consiguiente, el procesado de los antígenos clase II es sensible a la inhibición tanto por agentes lisosomotrópicos -cloroquina y cloruro amónico que elevan el pH de estos compartimientos- como por los inhibidores proteásicos endosomales. Las moléculas clase I derivan sus péptidos de fuentes intracelulares y el proceso no es bloqueado por los anteriores agentes. Sin embargo, el procesamiento antigénico de la clase I, pero no la clase II, se inhibe por brefeldina A que bloquea el movimiento de las proteínas de membrana de ER al Golgi; y, en segundo lugar, por el producto del gen E3/19K adenoviral que se une específicamente a las moléculas de la clase I y las retiene en el retículo endoplásmico.
Varias líneas de evidencia apoyan la hipótesis de la influencia de los genes del complejo MHC sobre la susceptibilidad a diversas enfermedades con características autoinmunes.
1. La frecuencia de ciertos alelos/haplotipos HLA en grupos de pacientes no relacionados.
2. La cosegregación de alelos HLA y enfermedad dentro de familias.
3. Los datos de modelos experimentales autoinmunes.
Entre otras, la diabetes dependiente de insulina y la esclerosis múltiple están bajo dependencia poligénica. La identificación de genes susceptibles adicionales está siendo objeto de un estudio permanente en la actualidad, si bien pocos genes distintos de los de la clase I y II han podido ligarse a la herencia de alguna enfermedad particular. Una excepción la constituye el lupus eritematoso sistémico que, además de su asociación a HLA-DR3, está asociado con ciertas formas alélicas de los genes del sistema de complemento.
Tras su clonación en 1984, el receptor de las células T (TCR) y sus genes, y ß, se han constituido en candidatos a incidir sobre la susceptibilidad de enfermedades asociadas a HLA. El primer requisito, el polimorfismo alélico, parece demostrarse en los análisis del polimorfismo de la longitud de los fragmentos de restricción (RFLPs). Algunos de estos esquemas alélicos se han asociado con distintas enfermedades-HLA; sin embargo, no existen aún datos consistentes que permitan atribuir las enfermedades autoinmunes a la herencia génica TCR.
1. VIRUS, CANCER Y HLA
En los últimos años está ganando terreno la participación de los productos del complejo NHC en las relaciones ya establecidas entre infección viral y modificación de los genes supresores de tumores. Entre las relaciones descritas se encuentran:
– La asociación entre el antígeno HLA-DR1 y la susceptibilidad a un carcinoma raro de tiroides.
– La conexión entre el antígeno HLA-DQw3 y el carcinoma cervical de células escamosas.
– La regresión y la conversión maligna de los papilomas virales de conejo se ha encontrado llevarse a cabo bajo control genético; la primera en relación con el fragmento DR EcoRI y la segunda asociada al fragmento DQ PvuII.
Otro tipo de relación recientemente descrita se refiere a las implicaciones de los marcadores génicos del MHC en la selección sexual en ratones. No deja de ser sugestiva a este respecto la hipótesis de tal discriminación por el olor de la orina, a su vez, relacionado con alelos MHC.
2. MIGRACIONES
En lo que respecta a la biología humana, el Rasgo genético fundamental de las migraciones es su capacidad de transferencia de genes desde una localización geográfica a otra. No hace falta insistir en toda la gran variedad de factores que aparecen involucrados, ya sea en las causas o en los efectos de las migraciones. y, como simple mención, no hay que olvidar las implicaciones sociales y económicas de estos movimientos, así como la gran variedad de su naturaleza: internacional o interna; local, urbana o Rural, etc. Desde el punto de vista biológico se acostumbra a distinguir las migraciones al azar de las que tienen lugar de forma selectiva. Distinción de gran importancia para interpretar las diferencias existentes entre las características biológicas de la población donadora de la migración y las de la población migrante algún tiempo después de su inserción en la población receptora.
Desde un punto de vista estrictamente biológico, las migraciones pueden originar cambios en las frecuencias génicas de la población, si la migración ha ido asociada a un flujo génico.
3. FLUJO GÉNICO
Diseminación o migración efectiva de genes por efecto del entrecruzamiento. Como consecuencia de él, ciertas variantes genéticas pueden introducirse de una población a otra.
4. DERIVA GÉNICA
VCambios en la frecuencia alélica que resultan del muestreo al azar, de tamaño limitado, de los gametos en sucesivas generaciones.
FRECUENCIA ALELICA
Proporción de alelos de un determinado tipo en una población.
GAMETOS
Células sexuales haploides que poseen un sólo juego de cromosomas.
ALELOS
Formas particulares de un gen.
En lo que respecta a la biología humana se dan varias posibilidades de líneas de estudio. En cuanto las migraciones humanas constituyen un mecanismo que inserta DNA de un pul génico en otro, está claro que la migración marital, al igual que la pre- o post-marital, puede alterar la composición del patrimonio génico de la población receptora. Este tipo de aproximación experimental biológica a la migración como fenómeno transportador de genes, lleva consigo la dificultad de la determinación del genotipo de los individuos no migrantes, de los receptores y de la población resultante, con relación a ciertas características de interés.
5. MIGRACIONES HUMANAS
1. Mecanismo que inserta DNA de un pulgénico en otro.
2. Mecanismo que inserta una clase similar de individuos en ambientes diversos.
3. Mecanismo capaz de producir una interacción entre las influencias genéticas y las ambientales.
En segundo lugar, la comparación de los genotipos de los emigrantes -población resultante de su inserción en la población receptora- con los de aquellos no-emigrantes, puede suministrar datos estadísticos acerca de la influencia de los factores que difieren entre los entornos de los lugares donantes de la migración y los entornos de los lugares receptores de los inmigrantes. Los problemas de esta clase de estudios radican en la determinación de los factores que, dentro de un complejo sistema de variables, son los responsables de las diferencias biológicas encontradas.
En tercer término, para el estudio de las características genéticas de las migraciones es necesario confinar la atención a rasgos auténticamente hereditarios, como lo son los grupos sanguíneos, las proteínas séricas y los antígenos de los linfocitos humanos HLA. De otro lado, en el estudio de los efectos ambientales hay que seleccionar variables de naturaleza antropométrica o fisiológica. El área principal de interacción entre las variables génicas y las ambientales se encuentra en relación con las enfermedades humanas de etiología mixta. Algunos de los ejemplos más importantes de esta interacción son las talasemias, la anemia falciforme y la deficiencia de glucosa 6-P deshidrogenasa. El caso de la anemia falciforme es de singular facilidad en el estudio de tales interacciones por la influencia que sobre los portadores de las células patológicas ejerce el que los emigrantes procedan de localizaciones geográficas con ambiente de malaria o no lo sean.
Así pues, el rasgo genético fundamental de las migraciones es su capacidad de transferencia de genes de una localización geográfica a otra. Los emigrantes portan sus genes al territorio habitado por una población receptora; genes que se entremezclan con el pul génico de la población huésped. Con ello se produce simultáneamente: a) un incremento de la variabilidad genética dentro de cada una de ambas poblaciones, emigrante y receptora; b) una reducción de las diferencias genéticas que pudieran existir entre ellas. Dada la existencia de estos dos tipos de poblaciones, se da: a) un incremento de la variabilidad intrapoblacional y b) una disminución de la variabilidad interpoblacional. La forma con arreglo a la cual se produce el acercamiento génico de ambas poblaciones se estudia mediante diferentes tipos de análisis estadístico.
TALASEMIA
Anomalía patológica hereditaria caracterizada por una deficiencia en la producción de alguna de las cadenas, ( -talasemia) o ß (ß-talasemia), de la hemoglobina.
ANEMIA FALCIFORME
Alteración patológica hereditaria caracterizada por la mutación Glu->Val en la posición 6 de la cadena ß en la hemoglobina adulta. La desoxigenación de esta hemoglobina da lugar a la formación de geles y las consiguientes anomalías circulatorias.
GLUCOSA 6-P DESHIDROGENASA
Enzima que cataliza la oxidación directa de la glucosa y su conexión con la vía de las pentosas. La deficiencia de esta actividad en los eritrocitos provoca un tipo de anemia hemolítica que se exalta con la ingestión de ciertos medicamentos -antimaláricos del tipo de la primaquina, acetanilida, sulfanilamidas, nitrofurantoína- en los individuos sensibles. Alteración patológica hereditaria que afecta preferentemente a los individuos de la raza negra.
HEMOGLOBINA
Proteína transportadora de oxígeno en los eritrocitos. posee una estructura cuaternaria tetramérica, del tipo 2 ß2.
Lo más importante es que las migraciones son el vehículo de uno de los mecanismos mayoritarios de evolución. Evolución definida como cambio en la frecuencia génica de una población.
Así pues, los efectos genéticos de la migración lo serán sobre el pul génico de la población receptora y sobre el pul génico de la población de origen. Efectos genéticos que dependerán de la magnitud y de la naturaleza de las migraciones.
Uno de los ejemplos más clásicos de esta influencia es el de la población caribeña de Centroamérica. A comienzos del siglo XVII el pul génico de las Antillas menores estaba formado exclusivamente por genes amerindios. Entre 1517 y 1646 se añadió a este pul génico el componente africano, procedente de los esclavos y de los fugados o cautivos de los territorios limítrofes. Tras una serie de vicisitudes, una parte de la población, conocida como la de los negros del Caribe, se instaló en las Honduras británicas, Guatemala y Nicaragua, A partir de ello tuvo lugar una dispersión rápida a lo largo de unos mil kilómetros de la costa de América Central, con la subsiguiente entrada de genes europeos.
De esta manera, en la población resultante se ha podido detectar la contribución africana por la presencia de marcadores tales como el gen Su del sistema de grupos sanguíneos MNS, el haplotipo Gmz, a; b en los alotipos de inmunoglobulinas o el de la hemoglobina S. La presencia europea por la presencia del haplotipo Gmf;b del sistema Gm. La contribución de las comunidades indígenas se ha atestiguado por la presencia de los genes del grupo sanguíneo Diego y de la albúmina México. Con el uso de estos marcadores se ha llegado a la obtención de los siguientes valores como contribución relativa al acervo génico actual: africana, 60-80 por ciento; europea, hasta 16 por ciento; amerindia, 17-40 por ciento. Ejemplo de cómo los genes de una población inmigrante han llegado a predominar sobre los da la población receptora. Este mismo ejemplo de la población caribeña negra sirve para ilustrar cómo los genes portados por los emigrantes han alterado la eficacia selectiva de la población original de la región.
Se trata, como es bien conocido, de una región infestada por el mosquito Anopheles, portador de los parásitos de la malaria, principalmente Plasmodium vivax y Plasmodium falciparum. Y en el pul génico africano han evolucionado una serie de variantes génicas que otorgan al individuo protección frente a estas especies productoras de la enfermedad. Entre los sistemas que otorgan esta ventaja se encuentran los genotipos de hemoglobinas AS y AC y el homocigótico FyFy del sistema de grupos sanguíneos de Duffy. Estos genotipos no se encontraban entre las poblaciones indígenas ocupantes primitivas del territorio, ni tampoco en las de los europeos. Este tipo de transferencia génica ha significado, pues, un cierto éxito evolutivo en la colonización de estas zonas por los negros africanos.
HOMOCIGÓTICO
Individuo que posee alelos idénticos en segmentos cromosómicos homólogos.
HETEROCIGÓTICO
Individuo que posee alelos diferentes en segmentos cromosómicos homólogos.
GRUPO SANGUÍNEO
Cada uno de los tipos que resultan de la clasificación de la sangre de un individuo haciendo uso de los antígenos presentes en la superficie de los eritrocitos.
INMUNOGLOBULINA
Cada una de las proteínas que ejercen la función de anticuerpos. Constan de dos cadenas pesadas unidas entre sí por puentes disulfuro; cada una de las cuales se une, a su vez, de igual manera a una cadena ligera. Según la naturaleza de las cadenas pesadas resultan cinco tipos de inmunoglobulinas: A,D,E,G y M.
6. MIGRACIONES Y SALUD
Un aspecto muy importante de las migraciones se refiere a su relación con la diseminación de enfermedades y al cambio en los esquemas de susceptibilidad a las enfermedades. Tampoco es necesario acudir a los esquemas tradicionales de los movimientos de poblaciones; incluso en nuestro siglo las poblaciones se mueven por un gran número de razones. Entre ellas, las siguientes:
– Las migraciones temporales de los trabajadores agrícolas.
– La migración laboral más permanente, tanto intra- como inter-nacional. Son ejemplos, los movimientos de las poblaciones de las islas del pacífico hacia Australia y Nueva Zelanda; de América del Sur y Central hacia el Norte del Continente; de los habitantes del Norte de Africa, Yemen, Turquía, India, Pakistan, etc. hacia el occidente europeo.
– Las persecuciones políticas y religiosas de nuestros días.
– El turismo de corta duración.
Sin entrar en detalles particulares de cada uno de estos movimientos de poblaciones, puede concluirse la existencia de fenómenos tales como:
– La propagación de enfermedades comunicables y a través de insectos vectores.
Bien conocida es la aportación española a la difusión de enfermedades en el Nuevo Mundo, en poblaciones que, carentes de resistencia natural, fueron diezmadas por las oleadas de epidemias. A la inversa, los hábitos de los amerindios contribuyeron a la extensión del cáncer de pulmón.
Son clásicas asimismo a este respecto las epidemias de cólera, viruela, tifus, fiebre amarilla, meningitos, poliomielitis, tracoma, etc. difundidas por portadores. A su lado, la difusión de la malaria ha sido el ejemplo más típico de la propagación de enfermedades por insectos vectores; aunque la eliminación masiva de los mosquitos por DDT en algunas zonas del mundo ha conseguido detener en cierta medida esta propagación.
– Las enfermedades gastrointestinales tienen como causa fundamental las nuevas conductas alimentarias, abundantes en grasas e hidratos de carbono y responsables de manifestaciones patológicas del tipo de diverticulitis, hernias de hiato, cálculos biliares, etc.
– La arterosclerosis y las enfermedades cardiovasculares ha permitido las observaciones siguientes: a) su presentación en Japón es la cuarta parte de la que muestra la población japonesa en los Estados Unidos; b) las lesiones vasculares intracraneales en Japón son 2-3 veces superiores al número de las de los japoneses en Estados Unidos.
Otras veces, sin embargo, los emigrantes portan consigo los riesgos que padecen en sus propios países. Así, la elevada incidencia de las alteraciones isquémicas en Israel es llevada consigo por los judios emigrantes a Europa y Estados Unidos y repercute, incluso, en los nacidos en el lugar de destino.
– Es bien conocida la relación de la enfermedad cancerosa con el elevado nivel de contaminación de aire, agua y alimentos, consecuencia del progreso industrial y tecnológico. Entre otros productos contaminantes se encuentran los boliclorobencenos, las dioxinas, el formaldehido, el asbesto, los insecticidas y plaguicidas, el plomo de las gasolinas, los aditivos de alimentos, etc.
Por lo que se refiere al padecimiento de esta enfermedad por las poblaciones emigrantes se dan dos circunstancias principales: a) Entre los emigrantes la frecuencia es intermedia entre la de los sedentes y la de la población huésped. b) Los factores económico-sociales se encuentran asociados a la localización y frecuencia del cáncer: la frecuencia se eleva en aquellas localizaciones asociadas al mayor estándar socioeconómico, tales como pulmón,vesícula y colorectal; en tanto que disminuye la frecuencia en las localizaciones asociadas a un inferior estándar socioeconómico, tales como los cánceres de estómago y de cervix.
Si la migración supone una elevación del estándar socioeconómico, habrá que suponer el incremento de la enfermedad vinculada a la mejor calidad de vida.
Es un hecho, además, por lo general admitido, que las migraciones ejercen sobre sus componentes un cambio en los esquemas de susceptibilidad a las enfermedades. En cualquier caso, los movimientos de población desde el campo a la ciudad representan una modificación drástica en el entorno físico, cultural y social. Y muchos estudios recientes prueban que los emigrantes de zonas rurales a urbanas exhiben mayores riesgos de padecimiento de enfermedades que las propias poblaciones urbanas. Así, muestran mayor riesgo de enfermedades infecciosas -tuberculosis, por ejemplo- hipertensión y alteraciones coronarias, diabetes y gota.
Sin embargo, por otro lado, tambien parece demostrado que, al menos desde la década de los años 50, los emigrantes urbanos exhiben menor mortalidad a todas las edades que los sedentes rurales. Naturalmente que sobre todo ello ejerce una influencia notable el contexto socioeconómico entre las partes en que la migración actúa.
No tiene nada de extraño, pues, que los programas modernos de salud pública, los tratamientos médicos, el mayor nivel de educación y escolarización de los jóvenes e, incluso, la misma adaptación a las enfermedades de la urbanización, logren un mejor estado sanitario en los medios urbanos frente a la situación rural con mayores tasas de morbilidad y mortalidad.
Son muy numerosos los estudios realizados en esta dirección. Durante los primeros años de la década de los años 60 se realizó un amplio estudio comparado de la presión sanguínea en las mujeres zulues de Africa del Sur, en zonas urbanas y rurales. Las mujeres urbanas tenían un comportamiento hipertensivo superior al de las que seguían habitando las zonas rurales. Sin embargo, las residentes durante largo tiempo en las zonas urbanas veían disminuir este parámetro frente a las que aún residían en dichas zonas durante corto tiempo. Los autores del estudio epidemiológico han relacionado este comportamiento con el estrés emocional de la emigración. También hay que señalar que los individuos más susceptibles a la hipertensión son aquellos que no se logran integrar en la vida urbana y conservan sus hábitos y prácticas culturales rurales.
En las migraciones internas de los Estados Unïdos de América del Norte se citan datos epidemiológicos que demuestran una mayor prevalencia en las zonas urbanas de enfermedades cardiovasculares, frente a su presencia en las zonas rurales. En este mismo sentido, cuando la población rural de Carolina del Norte fue masivamente urbanizada por la influencia de la industria -el famoso Triangle park-, las enfermedades cardiovasculares de la población sedente se elevaron de manera extraordinaria ante el cambio en el comportamiento de vida.
Estos planteamientos conceptuales de índole general referidos a las migraciones y las relaciones encontradas entre migración y salud, destacan el interés epidemiológico de estos fenómenos sociales. Interés e importancia que aumentan de forma extraordinaria con la asociación establecida entre el riesgo de padecimiento de enfermedades autoinmunes y la posesión de ciertos marcadores polimórficos, propios del complejo MHC. Resulta obvio, pués, que de la naturaleza de las asociaciones y de los estudios poblacionales amplios acerca de la presencia de estos marcadores, se han de deducir datos fundamentales a los aspectos científicos y sociales de la medicina preventiva. Los rasgos biológicos de la diversidad de los individuos, que se han utilizado en los estudios genéticos de las poblaciones, aparecen en la tabla 4.
7. GENÉTICA DE ACONTECIMIENTOS PREHISTÓRICOS
Utilizando distintos tipos de marcadores biológicos se han obtenido datos acerca de la génesis de acontecimientos prehistóricos. Así, mediante las frecuencias génicas de seis loci de rasgos serológicos de los grupos aleutianos, esquimales y atabascanos, de Alaska, se ha podido construir una especie de reloj genético de las sucesivas bifurcaciones de la población nativa de América del Norte, sobre la base de trece grupos:
8 tribus atabascanas
1 tribu aleutiana
3 esquimales Yupik-hablantes
1 esquimal Inupiak
Con el empleo de las frecuencias génicas de estos grupos, publicadas en 1979, se ha podido calcular la divergencia entre grupos y la divergencia dentro de un grupo. Suponiendo que las fuerzas evolutivas que operan sobre cada locus son relativamente iguales para todas las poblaciones en largos periodos de tiempo, las relaciones de distancias entre grupos son proporcionales al intervalo de tiempo desde la bifurcación de cada grupo y las relaciones de diferencias dentro de cada grupo son proporcionales al tiempo en que cada población ha acumulado la variación genética desde su origen. Por tanto, la diversidad interna de cada grupo se iguala en el origen y la diversidad entre grupos se iguala en la bifurcación o divergencia. Las relaciones se convierten en unidades temporales por fijación de una fecha de partida. Así, se ha fijado hace nueve mil años la fecha de divergencia inicial de esquimales y aleutianos, utilizando como dato experimental el análisis de radiocarbono en Alaska y Umnak, respectivamente.
De esta manera resulta que los indios atabascanos y los mongoloides del mar de Bering se bifurcaron hace 15.000 años; los aleutianos y los esquimales se separaron hace 9.000 años; y los esquimales Inupiaky Yupiak lo hicieron hace 5.000 años. Un dato de cierto interés resulta de la confrontación de estos resultados genéticos con las estimaciones cronológicas de diferenciació linguística. De tal comparación resulta que la divergencia de las poblaciones precede a la diferenciación lingüistica.
8. UTILIZACIÓN DEL SISTEMA HLA
La complicada realización experimental del análisis del complejo HLA tiene a su favor el ser extraordinariamente informativo.
En 1984 se llevó a cabo un análisis de la distribución de la frecuencia génica de HLA en diecisiete poblaciones del pacífico. Entre otros datos se ha encontrado que los aborígenes australianos no pueden agruparse con las poblaciones de Melanesia, Micronesia y Polinesia, debido a que los australianos presentan un intervalo restringido de antígenos HLA. Sin embargo, el perfil australoide de HLA es compartido por poblaciones papues de Nueva Guinea. Australia y Nueva Guinea debieron mantener contacto hasta unos diez mil años, puesto que el ligamiento A11.B40 persiste hoy en ambas poblaciones. Estos datos genéticos se están interpretando en la actualidad a la luz de los obtenidos previamente en las investigaciones paleontológicas, arqueológicas y antropológicas.
Hace cuatro años se obtuvo el árbol familiar de los orígenes del hombre basado en los mapas de restricción del DNA mitocondrial de unos 140 individuos de diferente asignación racial y localización geográfica diversa.
La conclusión fundamental de estos estudios ha sido el origen ancestral común del genoma mitocondrial humano, localizado en una mujer, de hace 200.000 años del Africa subsahariana. Conclusión que ha sido objeto de múltiples evocaciones populares como Eva del Edén africano.
Estudios más recientes prueban que si bien todo el DNA mitocondrial presente en la población humana actual se originó de una mujer, hace 200.000 años, no se sigue de ello que esta mujer fuera la madre común habida cuenta de los miles de genes implicados. Estos genes pueden rastrearse hasta llegar a los diversos padres y madres que vivieron en diferentes lugares y diferentes tiempos pasados. Los datos del DNA mitocondrial no permiten, pués, hacer conjeturas sobre el tamaño y la composición de la población original.
MITOCONDRIAS
Orgánulos propios de las células eucarióticas. Poseen DNA y una serie de sistemas enzimáticos, entre los que destacan los responsables de la respiración celular.
DNA MITOCONDRIAL
Genoma presente en las mitocondrias. En el hombre responde a la estructura de doble hélice circular cerrada, con unos 16.500 pares de bases.
El ordenamiento de los genes mitocondriales responde en los vertebrados a un elevado grado de conservación.
De otro lado, la secuencia de bases en el DNA mitocondrial ha evolucionado 5-10 veces más rápidamente que la del DNA nuclear.
Debido a estas propiedades y a la relativa facilidad de su obtención, el DNA mitocondrial es una molécula clave en los estudios de la evolución y la genética de poblaciones.
Las moléculas de DNA mitocondrial se heredan vía materna.
El polimorfismo apuntado en las clases, familias, subclases y loci del complejo principal de histocompatibilidad (MHC) (tabla 5) es único en su amplitud y en la distancia genética entre alelos individuales. Entre 6 y 57 alelos se han identificado en los diferentes loci polimórficos; presente cada alelo en las poblaciones humanas a frecuencias apreciables. El número de alelos, al menos en ciertos loci, se presume superior a 100. También es un hecho notable que la diferencia entre alelos puede afectar hasta 90 o más sustituciones nucleotídicas y que sus productos, los alomorfos, pueden llegar a diferir hasta 20-30 sustituciones de aminoácidos.
Artículo extraído del nº 33 de la revista en papel Telos